Karnataka 2nd PUC Biology Notes Chapter 2 Sexual Reproduction in Flowering Plants
The Flower: A fascinating organ of Angiosperms
Flower is the modified shoot of an angiosperm meant for sexual reproduction. Vegetative bud present at the axil of the leaf or at the apex of stem undergoes physiological changes and transforms into flower bud during the reproductive phase of the plant.
Parts of a typical flower:
A typical dicot flower shows the following parts:
1. Pedicel: It is the stalk of the flower by which flower is attached to the reproductive shoot.
2. Thalamus: It is the bulged tip of the pedicel on which floral whorls like calyx, corolla, androecium and gynoecium are present.
![]()
3. Calyx: It is the outermost whorl of thfe flower. It is small green and consists of a number of sepals.
4. Corolla: It is the second inner whorl of the flower. It is comparatively large, brightly coloured and consists of a number of petals.
5. Androecium: It is the third and male reproductive whorl of the flower. It consists of a number of stamens.
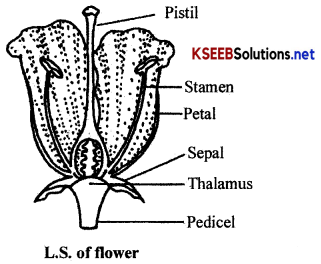
6. Gynoecium or Pistil: Itis the innermost and female reproductive unit of the flower. It is composed of one to many carpels.
Among the four floral whorls of the flower, calyx and corolla are regarded as accessory whorls, as they serve secondary purposes (serve as protective structures of sex organs and attraction of insects for cross pollination), whereas androecium and gynoecium are regarded as essential whorls, as they serve as sex organs and essential for sexual reproduction.
Pre-Fertilisation: Structures and Events
1. Stamen, Microsporangium and pollen grain: A typical stamen consists of a long and slender stalk called filament and the terminal generally bilobed structure called the anther.
The number and length of stamens are variable in flowers of different species.
A typical angiosperm anther is bilobed with each lobe having two theca (chambers) i.e. they are deithecous. The anther is a four sided tetragonal structure consisting of four microsporangia located at the corners, two in each lobe.
The microsporangia develop further and become pollensacs which extend all through the length of an anther and are packed with pollen grains.
(a) Structure of a mature anther (Microsporangium): Mature anther internally contains four microsporangia, two in each anther lobe. Each microsporangium consists of an outer anther wall and inner anther locule
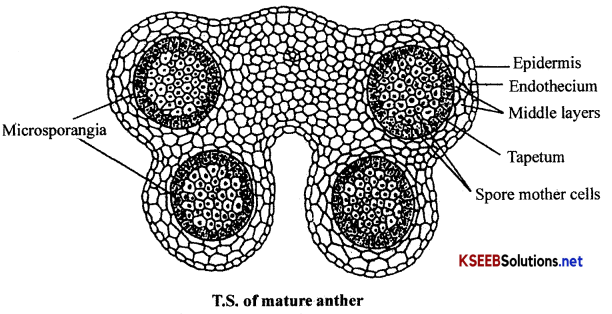
(i) Anther wall: It is the protective covering of anther or microsporangium. It is composed of epidermis, endothecium, middle layer and tapetum.
1. Epidermis: It is the outermost protective layer of anther composed of a single layer of cells. The cells are often stretched and flattened in mature anther.
2. Endothecium: It is the uniseriate layer composed of radially elongated cells with callose thickening on their inner tangential walls. Along the line of dehiscence of anther lobe, endothecium is composed of thin walled cells called stomium.
3. Middle layers: These are 2-3 concentric layers of cells present in between the endothecium and tapetum. In mature anther the cells of middle layers are flattened and collapsed due to the pressure exerted in the anther locule.
4. Tapetum: This is the innermost layer of anther wall composed of secretory cells. These cells secrete nutrients for developing microspore mother cells/ pollen grains in the anther locule.
![]()
Microsporogenesis (Formation of pollen grains): The process of formation of microspores from a pollen mother cell (PMC) through meiosis is called microsporogenesis. Each cell of sporogenous tissue is capable of giving rise to a microspore tetrad, hence acts as, potential pollen mother cell. The microspores are arranged in a cluster of four cells, called microspore tetrad. As the anthers mature and dehydrate, the microspore, dissociate from each other and develop into pollen grains. Inside each microsporangium, several thousands of microspores and pollen grains are formed.
(ii) Anther locule: It is the sporangial cavity containing a number of microspore tetrads / pollen grains formed from microspore mother cells by meiosis. (Microspores after the separation from tetrad are called pollen grains)
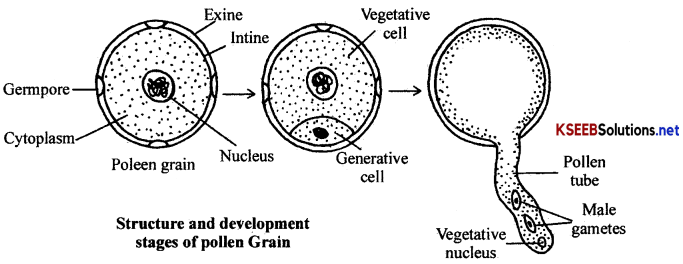
(b) Structure of pollen grain: Pollen gram is the haploid unicellular, uninucleated spherical shaped body bounded by outer thick, spiny or reticulate callose wall called exine and inner thin, smooth cellulose wall called intine. In the exine, thin areas called germ pores are present. Intine encloses peripheral cytoplasm and central nucleus.
Pollen grain represents the first stage of male gametophyte. Development by pollen grain takes place when it is still present in the anther locule. During the development it divides unequally into a large vegetative cell and a small generative cell. Generative cell then divides and give rise to two non-motile male gametes. This takes place either in the pollen grain or in the pollen tube after pollination.
1. Pollen grains of many species cause severe allergies and chronic respiratory disorders like asthma, bronchitis etc. e.g. parthenium (carrot grass)
2. Pollengrains are rich in nutrients. Thus pollen tablets are used as nutrient supplements.
Nice to know:
1. Pollen consumption has been claimed to increase the performance of athletes and race horses.
2. The period for which pollen grains remain viable is highly variable and depends on the prevailing temperature and humidity. In some cereals such as rice, wheat, pollen grains lose viability within 30 minutes and in some members of Rosacea, leguminosae and solanaceae they remain viable for months.
![]()
Gynoecium or pistil:
It is the female reproductive and essential whorl of a flower. It is composed of one to many carpels. Gynoceium or pistil has three parts: ovary, style and stigma: Internally, ovary contains one to many ovules on placental tissue.
Note: In some cas6s, a pistil never develops ovules, such sterile pistil is called pistillode.
Carpel: Carpel is the basic unit of pistil or gynoecium. A pistil with one carpel is called simple pistil and with more than one either free or fused carpels, is called compound pistil.
Note: Carpel represents megasporophyll.
Types of Gynoecium:
I. Based on number of carpels:
(a) Monocafpellary – e.g:: bean; pea (1 carpel)
(b) Bicarpellary – e.g.: brinjal, mustard (2 carpels)
(c) Tricarpellary – e.g.: onion, castor (3 carpels)
(d) Poly carpellary – e.g.: lemon (Many carpels)
II. Based on number of locules: (Chambers)
(a) Monolocular – 1 chamber e.g.: cucumber
(b) Bilocular – 2 locules e.g.: tomato
(c) Trilocular – 3 locules e.g. : castor
(d) Multilocular -many locules e.g.: lemon
III. Based on fused or free status:
(1) Apocarpous: When carpels are free e.g.: michelia
(2) Syncarpous: When carpels are fused e.g.: hibiscus
(c) Structure of Mature Anatropous Ovule: Ovule represents the megasporangium. Anatropous ovule is an inverted ovule (due to 180° turn) in which lunicle and micropyle lie close to each other. It shows the following parts.
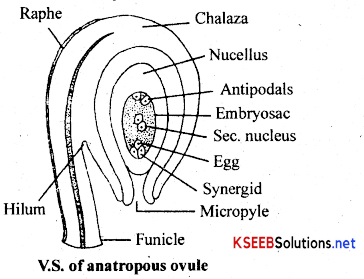
1. Funicle: It is the stalk of the ovule by which ovule is attached to placenta in the ovary.
2. Hilum: It is the region at which ovule is attached to the funicle.
3. Raphae: It is the extended part of the funicle beyond the hilum.
4. Chalaza: It is the basal part of the ovjule at which integuments originate.
5. Integuments: These are outer protective coverings of the ovule. These completely enclose the nucellus except for the region at the apex called micropyle.
6. Nucellus: It is the female gametophyte present in the nucleus towards the micropylar end. It consists of three antipodal cells towards chalazal end, secondary nucleus at the centre and egg appartatus towards the micropylar end. Egg apparatus consists of central egg cell and two lateral synergid cells.
7. Embryosac : It is the female gametophyte present in the nucleus towards the micropylar end. It consists of three antipodal cells towards chalazal end, secondary nucleus at the centre and egg appartatus towards the microphlar end. Egg apparatus consists of a central egg cell and two lateral synergid cells.
![]()
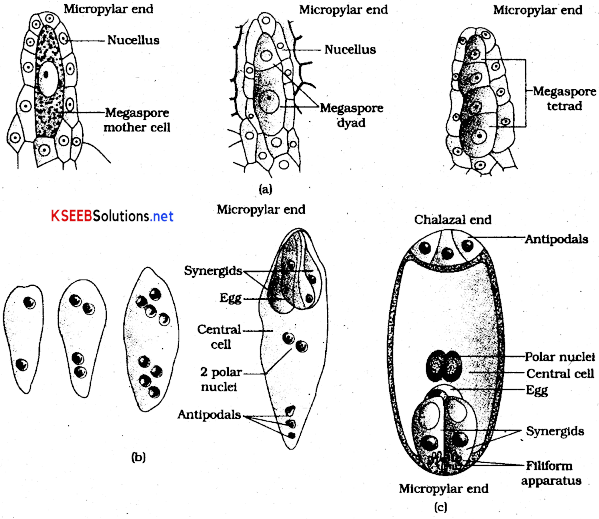
Megasporogenesis:
- A single megaspore mother cell is differentiated in the micropylar region of the nucellus of an ovule.
- This cell is larger and contains dense cytoplasm and a prominent nucleus.
- It undergoes meiosis and forms a cluster of four haploid cells, called megaspore tetrad.
- Of these, three degenerate soon and only one megaspore becomes functional.
Development of Female Gametophyte:
- The female gametophyte develops from a single megaspore and hence is described as monosporic development.
- The functional megaspore is the first cell of female gametophyte of angiosperms.
- It enlarges to form the female gametophyte, also called embryo sac.
- Its nucleus undergoes a mitotic division and the two nuclei move to the opposite poles, forming the 2-nucleate embryo sac.
- Two successive mitotic divisions in each of these two nuclei result in the formation of an 8- nucleate embryo sac.
- Cell wall formation starts at the eight-nucleus stage, resulting in the formation of a typical female gametophyte.
- Three nuclei are grouped together at the micropylar end to form the egg apparatus, consisting of two synergids and a female gamete/egg cell by wall formation.
- The remaining two nuclei are called polar nuclei, they move to the centre of the embryo sac and fuse to from a diploid secondary nucleus. The remaining three nuclei aggregate at the chalazal end to form three antipodal cells.
- Thus a typical angiosperm embryo sac is 8-nucleate and celled.
Pollination in Angiosperm:
In angiosperms, fertilization will be effective only when pollen grains from anther locule are transferred on to the stigma of gynoecium. Hence pollination is an important biological act. The transfer of pollen grains from the anther of a flower to the stigma of the same flower or another on the same plant or a different plant of same species or allied species is known a pollination.
Kinds of pollination: Pollination is of two kinds namely,
(i) self pollination and
(ii) cross pollination.
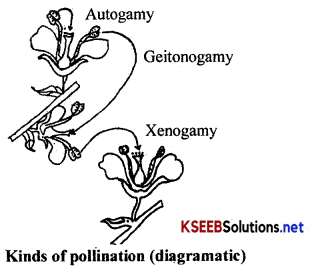
(i) Self pollination or autogamy: When the pol len grains pol linate the stigma of the same flower, it is called self-pollination.
(ii) Cross pollination or allogamy : When the pollen grains of one flower pollinate the stigma of the another flower located on the same plant or on the different plant of the same kind, it is called cross pollination. Cross pollination is of three kinds.
(1) Geitonogamy: Cross pollination which takes place between two flowers borne on the same plant is called Geitonogamy. From the genetical point of view there is a little difference between geitonogamy and autogamy.
(2) Xenogamy: Cross pollination which takes place between two flowers borne on two different plants of the same species is called xenogamy.
(3) Hybridisation: Cross pollination between the two flowers borne on two different species or genera is called hybridisation.
![]()
Contrivance for self pollination:
The following conditions or contrivance in the flowers ensure self pollination:
1. Cleistogamy: This is the condition where bisexual flower never open during anthesis (dehiscence of anther). Hence, self pollination is obligatory. Cleistogamic condition is seen in Balsam, oxalis.
2. Homogamy: This is the condition where carpels and stamens of a flower mature at the same time. This ensures self pollination. Homogamic condition is seen in Argemone, Mirabilis etc.
Contrivances for Cross Pollination: [Out breeding devices]
The following contrivances ensure cross pollination:
1. Dicliny: It is the condition where one of the two sexes is absent in the flower and flower becomes unisexual male or female (diclinous). Such diclinous flowers may be borne either on the same plant or on the two different plants Jn such cases, cross pollination is the rule. e.g.. maize, cucurbita.
2. Herkogamy: (Herkos = barrier) It is the condition where style of the gynoecium extend far beyond the anthers or stamens may face outward or pollens may aggregate into pollinia. In such cases, self pollination is impossible, e.g. gloriosa, calotropis.
3. Dichogamy: It is the condition where androecium and gynoecium in a flower mature at different times. In such a case, self pollination is ineffective, however it may take place at later stage if cross pol lination fails. Dichogamy may be of two types:
(a) Protandry: Anthers mature earlier than the carpels, e.g. sunflower, cotton.
(b) Protogyny: Carpels mature earlier than anthers, e.g. michelia, ficus.
4. Self sterility: It is the condition where pollen grains fail to germinate on the stigma of the same flower. In such cases, self pollination is ineffective and cross pollination is a must. e.g. tobacco, potato.
5. Heterostyly: It is the condition where the flowers on same plant have styles of different sizes, where one has short style and long stamens, while another has long style and short stamens. In such cases self pollination is impossible and cross pollination is a must, e.g. oxalis, primula.
Types of cross pollination: Cross pollination is always brought about by some external agents. Because pollen grains have no power of independent movement. Based on the agents involved in cross pollination, the following types of cross pollination are recognised.
1. Hydrophily: Cross pollination by the agency of water is called hydfophily and such flowers are called hydrophilous flowers. Hydrophily can be seen in some submerged hydrophytes (aquatic plant) like hydril la, elodea, vallisneria, etc.
Characters of Hydrophilous flowers:
- Generally, the pollen grains of submerged plants like hydrilla have same specific gravity as that of water, so that pollen grains suspend and float in water at different depths.
- In vallisneria special types of characters are developed for cross pollination. It is dioecious. Male plant bears small sessile male flowers in submerged spadix. These flowers detach and float on the surface of the water. Female plant bears solitary female flowers on the spirally coiled stalks.
- When flowers mature the stalks uncoil and bring the flowers to the surface of the water. When male flowers come in contact with a female flower, the anthers burst open and sticky pollen grains deposit on the stigma. After the pollination, the stalk of the female flower coils and thus flower is brought down into water, where fruit formation takes place.
2. Anemophily: Cross pollination by the agency of wind is called anemophily and such flowers are called anemophilous flowers. Anemophily can be seen in maize, grass and palms, etc.
Characters of anemophilous flowers:
- The flowers are small and do not have any colour or scent.
- Anthers are versatile, swinging freely in the air.
- Pollen grains are dry, light, smooth, powdery and produced in large quantity, so that they can be easily carried by wind.
- Stigmas may be feathery or long branched and bushy, so that they can catch the pollens from air easily.
![]()
3. Zoophily: Cross pollination by the agency of animals is called zoophily and such flowers are called zoophilous flowers.
Zoophily is again sub-divided into
(a) Entomophily – pollination by insects, e.g. sunflower, canna.
(b) Ornithophily -pollination by birds, e.g. silk cotton, passion flower,
(c) Chiropterophily – pollination by bats. e.g. bauhinia, banana.
Characters of Entamophilous Flowers: In majority of the plants, cross pollination takes place through agents like insects and such flowers are called entomophilous flowers. These show the following characters.
Colour: Normally petals of these flowers are brightly coloured to attract insects for the pupose to carryout cross pollination. In some plants, other parts of the flower are also modified into petaloid structure, e.g. In bougainvellia, bracts are brightly coloured, in canna stamens are petaloid.
Nector: It is the sugary substance secreted by gamopetalous corolla of entomophilous flowers. Insects while collecting nector, unknowingly bring-about cross pollination. In some plants, nectary glands are developed on involucre of cyathium inflorescence, e.g. poinsettia.
Scent: Some entomophilous flowers emit a strong scent during night, to attract the insects and take their service in cross pollination, e.g. nightqueen, jasmine.
Pollen Pistil interaction:
Recognition of compatible pollen:
- The stigma/pistil has the ability to recognize the right type of pollen i.e, the compatible pollen of the same species.
- The pistil rejects the pollen grains of other species and also the incompatible pollen grains of
the same species. - It is the result of interaction between the chemical components of the pollen and those of stigma.
Note: The inability of certain gametes even from genetically similar plant species, to fuse with each other is called incompatibility. It is also called intraspecific incompatibility, self sterility or self incompatibility.
Artificial Hybridisation:
It is one of the major approaches of crop improvement programme. It is a crossing exjperiment to make sure that only the desired pollen grains are used for pollination and the stigma is protected from contamination (from unwanted pollen). This is achieved by emasculation and bagging techniques.
Emasculation: If a plant bears bisexual flowers, removal of anthers from the flower bud before the anther dehisces using a pair of sterile forceps is necessary to avoid self pollination. This is referred as emasculation.
Bagging: Emasculated flowers have to be covered with a bag of suitable size, generally made up of butter paper to prevent contamination of its stigma with unwanted pollen. This process is called bagging.
When the stigma of bagged flower attains receptivity, mature pqllen grains collected from anthers of the male parent are dusted on the stigma, and the flowers are rebagged and the fruits allowed to develop.
Note: If the female parent produces unisexual flowers there is no need for emasculation. The female flower buds are bugged before the flowers open. When the stigma becomes receptive, pollination is carried out using the desired pollen and the flowers are rebagged.
![]()
Fertilization in Angiosperms:
Definition: Fusion of male and female gametes during sexual reproduction is known as fertilization. In angiosperms, two male gametes take part in fertilization, where one fuses with the egg and other with the secondary nucleus. Because of two fusion processes fertilization in angiosperms is referred to as double fertilization. S.G. Nawaschin first observed double fertilization in Fritillaria plant.
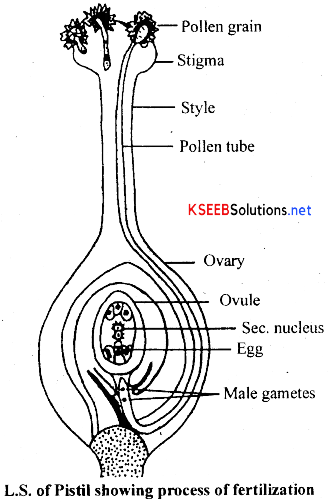
A brief account of double fertilization: The process of double fertilization can be explained under the following steps:
(i) Germination of pollen grains and growth of pollen tube:
- After pollination, pollen grain germinates, where intine of the pollen grain grow into a narrow tubular structure called pollen tube through the germ pore.
- Pollen tube thus formed carries most of the cytoplasm of vegetative cell, vegetative nucleus and two non-motile male gametes at the apex.
- Pollen tube grows through the stigmatic papillae, style, ovary wall and ultimately it reaches the ovule in the uvary.
(ii) Entry of pollen tube inside the ovule: Pollen tube enters the ovule either through the micropyle or chalaza or integuments. If pollen tube enters inside the ovule through the micropyle, it is called porogamy, if it enters through the ehalazal end, it is called chalazogamy and if it enters from lateral side through the integuments, it is called mesogamy.
(iii) Entry of pollen tube inside the embryosac: Irrespective of the entiy of pollen tube inside the ovule, it enters inside the embryosac through the micropylar end either between egg cell and one of the synergid cells or between embryosac wall and synergid cell or through the synergid cell. Then pollen tube discharges the male gametes through the terminal or sub-terminal pore developed in the pollen tube.
(iv) Double fertilization: One of the two male gametes fuses with the egg cell. This is known as syngamy and the fused product is called oospore. The other male gamete, unites with the secondary nucleus. This is known as triple fusion (as this involves the fusion of 1 male nucleus and 2 polar nuclei) and the fused product is called primary endosperm nucleus. (PEN).
Post fertilization changes in the flower: After fertilization the following changes take place.
- Oospore develops into embryo and PEN develop into endosperm.
- Ovule develops into seed and integuments of ovule develop into seed coat.
- The outer integument of the ovule forms the testa or outer seed coat.
- The inner integument forms the tegmen or inner seed coat.
- Ovary develops into fruit and ovary wall develops into fruit wall called pericarp.
- Style, stamens, calyx and corolla wither off.
![]()
Significance of double fertilization: Importance of double fertilization is the production of seeds from ovules and fruits from ovaries. Double fertilization also signifies the formation of nutritive tissue endosperm in the seeds.
1. Endosperm formation : The endosperm is developed from the triploid (3N) primary endosperm nucleus. The endosperm development precedes embryo development, because, it is a nutritive tissue that supplies food material to the growing embryo and also the seedling.
Soon after fertilization, the primary endosperm nucleus begins to divide and ultimately gives rise to the endosperm. The endosperm grows by absorbing the food supplied by the parent plant. Depending on the mode of formation, the angiospermic endosperm is of three types – free nuclear, cellular and helobial.
i. Free nuclear endosperm: The primary endosperm nucleus divides freely into large number of nuclei without any immediate cell wall formation. These nuclei are pushed towards the periphery and a large vacuole appears in the centre of the embryo sac. Later, the wall formation starts from periphery and progresses towards the centre and ultimately acellular endosperm is formed, e.g.: Cdpsella. jin certain cases, the cell wall. formation remains incomplete. For example, the coconut water (coconut milk) from tender coconut that you are familiar with, is nothing but free nuclear endosperm (made up of thousands of nuclei) and the surrounding white Kernel (coconut meat) is the cellular endosperm.
ii. Cellular endosperm: Each division of primary endosperm nucleus is followed by wall formation. Therefore, endosperm becomes cellular from the very beginning e.g. datura, petunia, balsam.
iii. Helobial endosperm: It is an intermediate type of endosperm formation between the nuclear and the cellular types of endosperms. The first division of the primary endosperm nucleus is followed by wall formation. As a result, two cells-micropylar and chalazal, are formed. Further development in both the cells occurs like that of nuclear endosperm. Helobial endosperms are generally found in monocotyledons.
Embryo
- The embryo formation starts after a certain amount of endosperm is formed, as there is an assured supply of nutrition to the embryo.
- The zygote divides by mitosis to from a proembryo first.
- Later development results in the formation of globular and heart-shaped embryo, that ultimately becomes the horse-shoe shaped mature embryo, with one or two cotyledons.
Development of the embryo: After fertilization, the diploid zygote secretes a wall around itself to become an oospore and divides into 2 cells. The upper cell which is away from the micropyle is called as embryonal cell and the lower cell towards the micropyle is called as the suspensor cell. The suspensor cell divides in one plane to produce a filament of cells called suspensor which elongates and thereby pushes the embryo deeper into the endosperm tissue for better nutrition. The basal cell of the suspensor is highly enlarged to form a food absorbing structure.
The embryonal cell produces a group of 4 cells called quadrant which further divides to give rise to an octant. These cells are the embryonal mass from which the future embryo develops. The cell of the suspensor adjacent to the embyonal mass is called hypophysis. It produces the apex of the radicle. The cells of the embryonal mass towards the suspensor produces the radicle. The embryonal cells away from the suspensor produce the 2 cotyledons (embryonic leaves) and plumule (the future shoot system).
The embryo passes through the globular stage and a heart shaped embryo state where the two cotyledons are clearly visible. Then it passes through the torpedo stage (called so because the elongated cotyledons look like a torpedo). During the embryo development the endosperm is partly or completely utilized.
![]()
Dicotyledonous Embryo:
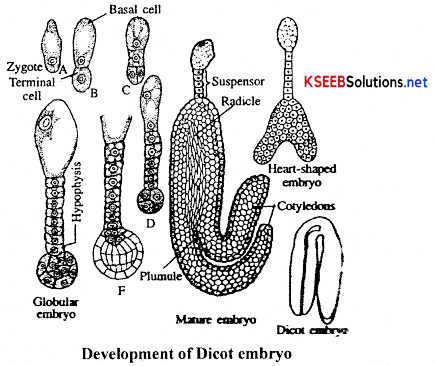
- In dicotyledonous plants, the embryo consists of two cotyledons and the embryonal axis between. them.
- The portion of embryonal axis above the level of attachment of cotyledons is the epicotyl and it terminates in the plumule (shoot meristem).
- The portion of embryonal axis below the level of attachment of cotyledons is the hypocotyl; It terminates in the radicle (root tip).
Monocotyledonous embryo (Grasses):
- In monocotyledonous plants like rice, maize, etc., the embryo has only one cotyledon (called scutellum) pushed towards one side of the embryonal axis.
- The embryonal axis has the radicle on its lower end (hypocotyl). The radicle is covered by an undifferentiated sheath called coleorhiza.
- At its upper end (epicotyl), the embryonal axis has plumule. It is covered by a hollow foliar sheath called coleoptile.
Seed:
As embryogenesis is taking place, the ovule transforms itself into a seed and the integuments develops into the tegmen and the outer integement into the testa.
Fertilised ovule is called seed. After fertilization, the integuments of the ovule develop into seed coat, fertilised egg into embryo and primary endosperm nucleus into endosperm.
Types of seeds:
Exalbuminous or non-endospermic seeds. These seeds do not have endosperm as the endosperm is fully utilized. Such seeds are called as non-endospermic. e.g. beans, mango, sunflower, tamarind etc.,
Albuminous or endospermic seeds: If the endosperm is partly utilized then the seeds are said to be endospermic. e.g. – castor, custard apple, four O’ clock plant etc.
Note: Perisperm: Occasionally, in some seeds such as black pepper and beet, remnants of nucellus are also persistent. This residual, persistent nucellus is the perisperm.
1. Structure of a typical Dicotyledonous seed:
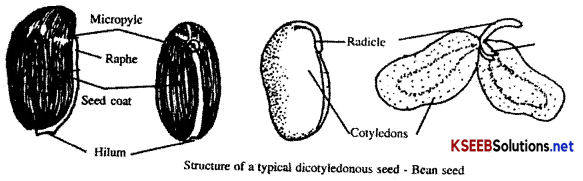
e.g.: Bean seed.
Bean seed is an exalbuminous seed or non endospermic seed, It is kidney shaped. It contains seed coat and embryo.
Seed Coat: It is the outer protective covering of the seed, which is brownish red in colour. It is differentiated into two layers namely, outer testa and inner tegmen. Testa is hard, brownish red layer developed from outer integument of the ovule. It is provided with white ridge called raphe and a broad scar called hilum. Micropyle is present as a small pore near the hilum. Tegmen is the thin white membranous layer which encloses the embryo.
Embryo: It consists of embryonal axis and a pair of fleshy cotyledons. The pointed end of the axis is called radicle and the leafy end of the axis lying between the two cotyledons is called plumule. The part of embryonal axis present just above the attachment of cotyledons is called epicotyl and part ofthe axis below the attachment is called hypocotyl.
![]()
2. Structure of a typical Monocotyledonous seed:
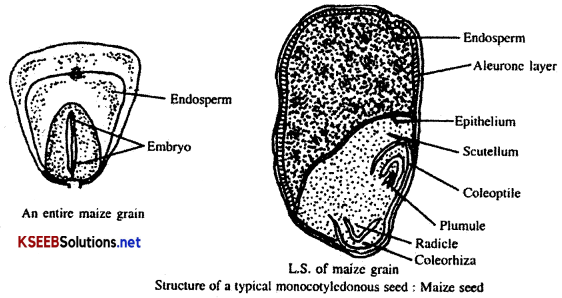
e.g.: Maize grain.
Maize grain is one seeded dry indehiscent fruit called caryopsis. It is an example for albuminous or endospermic seed. It has flat, oblong, coloured body and consists of grain wall called hull, endosperm and embryo.
Grain wall: It is the outer protective covering of the seed. It is yellow in colour. It consists of fruit wall (pericarp) and seed coat. These two layers are firmly attached and not separable from one another.
Endosperm: It forms the major bulk of the grain (2/3). It is composed of endospermal cells containing starch. It is externally bounded by a layer of cells called aleurone layer which contains mainly protein. Endosperm is separated from embryo by a layer of cells called epithelium.
Embryo: It occupies small area of the grain on one side of the base. It consists of a single shield shaped cotyledon called scutellum and embryonic axis. Embryonic axis consists of pointed ends called radicle and plumule. Radicle is protected by a sheath called coleorhiza and plumule by a sheath called coleoptile. At the time of germination, radicle develops into the root system and plumule into the shoot system.
Significance of seeds: Seeds offer the following advantages to the plants:
- Since reproductive processes such as pollination and fertilization are dependent on water, seed formation is more dependable.
- Seeds have adaptation for dispersal to new habitats and help the species to colonize in other areas.
- Seeds have enough food reserves to nourish seedlings until they are capable of photosynthesis on their own.
- The hard seed coat provides protection to the young embryo.
- Seeds are the product of sexual reproduction, hence they generate new genetic combinations leading to variations.
- Seed is the basis of our agriculture. Dehydration and dormancy of mature seeds are crucial for storage of seeds, which can be used as’ food through out the year and also raise crop in the next season.
Special mechanisms of reproduction:
(a) Apomixis :
1. It is a form of asexual reproduction, that mimics sexual reproduction, where seeds are – formed without fertilisation; it is also known as agamospermy.
2. Apomictic seeds may be formed in one or more of the following ways:
(a) A diploid egg cell (formed without meiosis during megasporogenesis) may develop in an embryo without fertilisation.
(b) Cells of the nucellus (diploid) surrounding the embryo sac may develop into embryos and become pushed into the embryo sac. e.g., Citrus, mango,
(b) Polyembryony:
1. It can arise due to one/more of the following reasons :
(a) More than one egg may be formed in the embryo sac.
(b) More than one embryo sac may be formed in an ovule.
(c) Other cells like synergids, or cells from nucellus may develop into embryos.
2. Polyembiyony is common is Citrus (orange, lemon), onion, mango, groundnut, etc.,
3. Polyembryony is more often associated with apomixis.
![]()
Fruits:
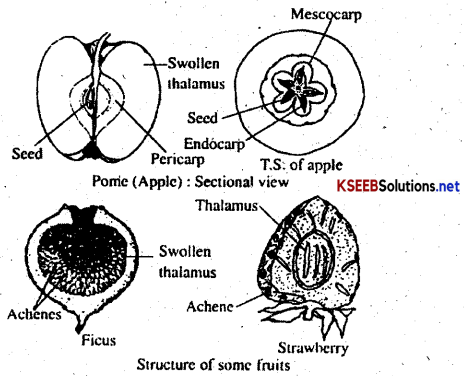
A fruit is defined as a fertilized and ripened ovary. The fruit wall is called pericarp which may be thin, thick or fleshly. The pericarp is differentiated into outer epicarp, middle mesocarp and inner endocarp. The fruit is said to be true when it is developed entirely from the ovary, e.g. mango. It is called a false fruit or pseudocarp, when any other floral part also contributes towards the formation of the fruit, e.g. apple, where the thalamus becomes fleshy and edible. Fruits that develop without fertilization are called parthenocarpic fruits. Such fruits are seedless since without fertilization ovules develop into seeds.
Significance of Fruits:
- Fruits provide protection to the seeds (till they mature), from the hostile environment and predators.
- They also help in the dispersal of seeds.